
生体電気と生命システム


生物学的機能を理解するための生体電気の役割について、ここ数年で重要な新しい発見がなされています。
生体電気は、生物の組織のあらゆるレベルで生物学的機能にとって重要な役割を果たしています。
The significance of bioelectricity on all levels of organization of an organism. Part 1: From the subcellular level to cells
Prog Biophys Mol Biol. 2023 Jan:177:185-201. doi: 10.1016/j.pbiomolbio.2022.12.002
電荷と原子

電子 (マイナス)と陽子 (プラス)の形の電荷は、生物システムのあらゆる場所に存在します。
カルシウム (Ca 2+ )、カリウム (K + )、ナトリウム (Na + )、鉄(Fe2+および Fe 3+)など、異なる電荷強度の正 (カチオン) および負 (アニオン)に帯電した原子 (イオン)も生物中に遍在しています。
これらの種類の帯電した物質は、電場 (静止時)、磁場 (一定速度で移動するとき)、および電磁場 (速度が変化するとき)を生成します。
生物学的システムでは、電流の流れはさまざまな構造に沿って、さまざまなメカニズムで発生します。
異なる符号と強さを持つ多種多様な種類の電荷は、電荷輸送、近距離および長距離の電気的相互作用、電力場、電磁放射など、生物学的組織の基本レベルにある多様な電気現象とプロセスの複雑な状況を作り出します。
生命は、電荷分離を引き起こすプロセスによって特徴付けられ、さまざまな構造レベルでのコンパートメントの複雑な入れ子構造をとります。
例えば、細胞質ゾルのイオン組成は細胞外環境とは異なります。K +の濃度は Na +の濃度より約 10 倍高く、細胞外空間ではその逆が当てはまります。
細胞内外の膜電位については、関連記事をご参照ください ↓
血液のゼータ電位については、関連記事をご参照ください ↓
分子と高分子

ニコチンアミドアデニン ジヌクレオチド(NAD + /NADH)やフラビン アデニン ジヌクレオチド(FAD/FADH2 )などの特定の分子が電荷担体として機能し、酸化還元反応に関与します。
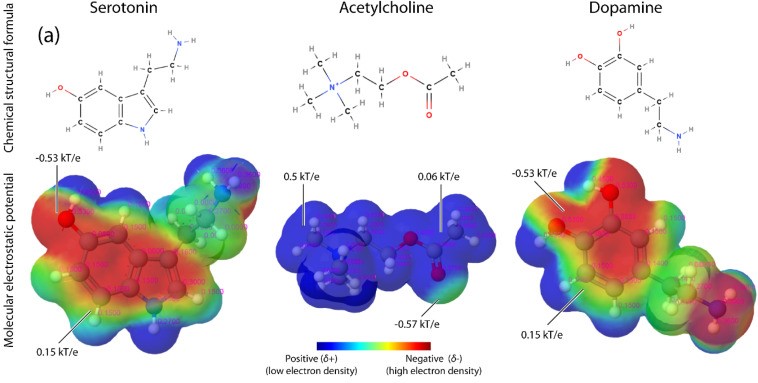
分子と高分子は、特定の電荷分布、すなわち静電ポテンシャル (Molecular Electrostatic Potential :MEP)を持っています。
一般的に電位は分子全体に均一に分布せず、電気双極子モーメントが生じます。
MEP は、位置に依存する静電引力と、反発力による分子間の相互作用において重要な役割を果たします。
生体分子の固有 MEP に起因する電気力は、タンパク質間相互作用、酵素-基質相互作用、タンパク質-核酸相互作用、またはタンパク質-脂質相互作用など多くの分子相互作用を支配します。
微小管とアクチンフィラメント
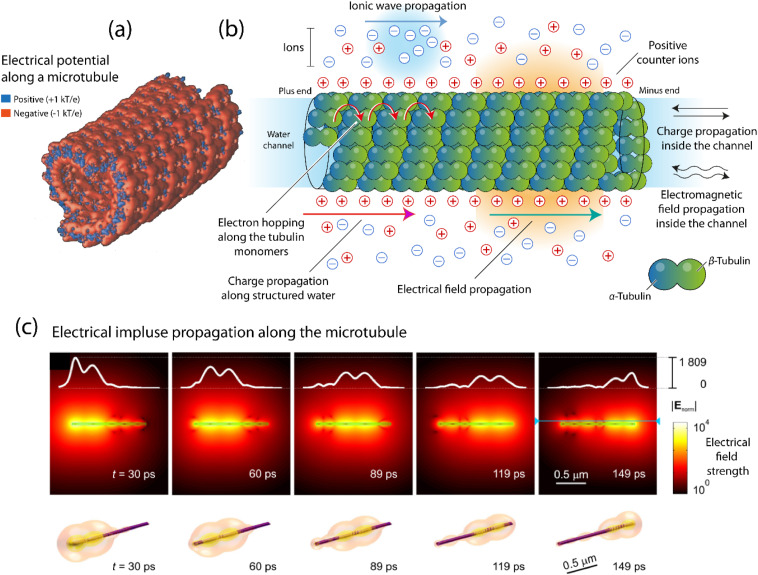
細胞内部では、細胞骨格(アクチンフィラメント、中間フィラメント、微小管で構成される) は、電荷とシグナル伝達を促進する構造ネットワークとみなすことができます。
微小管は、チューブリン二量体を含む円筒状のタンパク質ポリマーであり、その特有の電気的特性を持っています。
微小管に沿った電位、微小管内外の電荷と場の伝播など、微小管はさまざまな電気的側面を持っており、細胞内および細胞間の電気シグナル伝達に使用されています。
微小管とは別に、アクチンにも特有の生体電気特性があります。
F-アクチンは比較的高い表面電荷を持ち、正味負に帯電しています。
アクチン フィラメント (F-アクチン)は、繊維に沿った電荷の伝達を可能にするケーブルのような特性を持っています。
F-アクチン線維は軸索の内側にあるため、微小管を介した電気信号伝達に加えて、F-アクチンベースのイオン波や電気インパルスを介した脳細胞間の電気信号伝達の可能性があります。
つまり、細胞骨格全体が情報交換と処理のための電気ネットワークとして機能しています。
ミトコンドリアのエネルギー変換による微小管の電気振動については、関連記事をご参照ください ↓
核酸分子とタンパク質
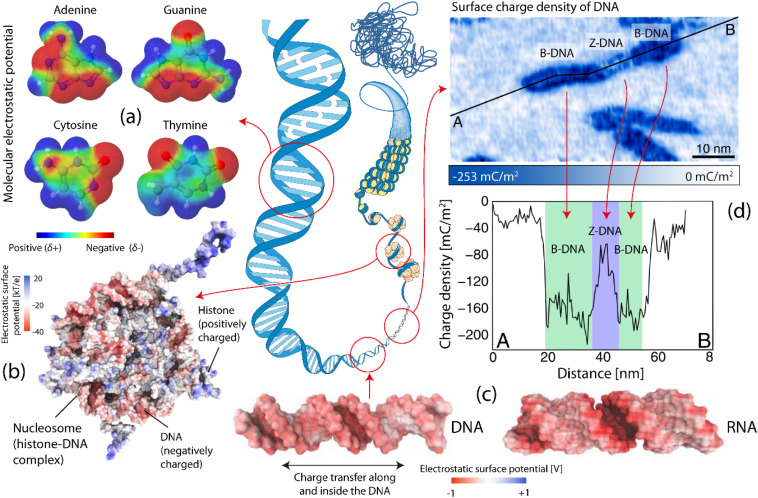
核酸分子は特定の電気的特性を持っています。
DNA の表面電荷密度はその立体構造によって異なります (Z 立体構造と比較して、 B 立体構造の電荷密度が高くなります)。
電気的特性は、DNA/RNA の構造安定性と立体構造に関連し、分子との相互作用を起こします。
二重らせん DNA 内の塩基対のスタックは、効率的な電荷輸送を促進します。
リボソームは非常にネガティブな MEP を持ち、リボソームの出口トンネルは特定の電気的機能を持っています。
トンネルの幅は 0.1 ~ 0.2 nmで水を含んでおり、マイナスに帯電しています。
リボソームによって組み立てられたタンパク質は、リボソームの出口トンネルを通過する必要があり、電位はリボソームでのタンパク質合成に重要な役割を果たしています。
新生ペプチドはトンネルと電気的に相互作用し、リボソームとペプチドの機能に影響を与え、ペプチドの電荷の再分配、翻訳速度の調節、シャペロン相互作用またはペプチドの折り畳みを引き起こす。
自然界では、大部分のタンパク質は中程度の正味電荷を持っていますが、一部のタンパク質は高度に帯電しています(過荷電タンパク質)。
タンパク質の電荷は周囲の溶液の pH に依存します。
タンパク質の電荷は、カチオン性 (リジン、アルギニン、ヒスチジン)およびアニオン性 (グルタミン酸、アスパラギン酸)アミノ酸残基によって均一に分布しません。
細胞小器官

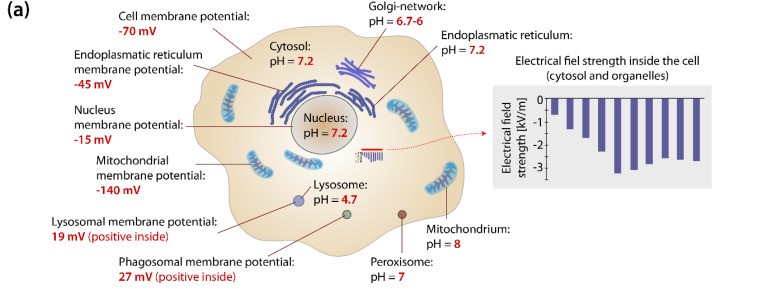
すべての細胞小器官は特定の膜電位 および pH 値 を持っています。生体電気によって複雑な細胞内環境を引き起こします。
膜電位については、関連記事をご参照ください ↓
pHについては、関連記事をご参照ください ↓
生物学的機能に対する電荷キャリアの流れの重要性は、ミトコンドリア内の電子伝達鎖(ECT) において明らかとなっています。
ミトコンドリア膜内のタンパク質複合体は、電子供与体と受容体の助けを借りて、電子とプロトンの共役移動を可能にしています。
電気化学的プロトン勾配を生成して、アデノシン三リン酸(ATP)の合成を可能にします。
ミトコンドリアのエネルギー代謝については、関連記事をご参照ください ↓
ミトコンドリアの電子伝達系については、関連記事をご参照ください ↓
ミトコンドリア
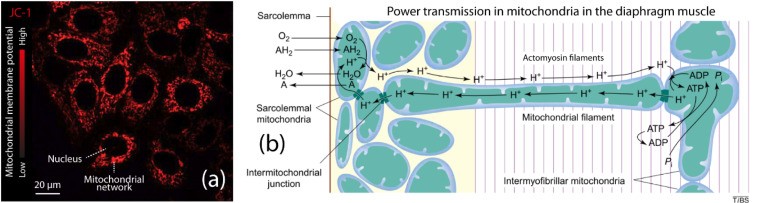
ミトコンドリアは、膜電位が高いだけでなく、電気的に結合できるミトコンドリア ネットワークを形成できます。
ミトコンドリアは電気伝達線維とみなすことができ、少なくとも 3 つの電荷伝達プロセスが起こる可能性があります。
1)K +、Cl –、Na +のようなイオン
2)ミトコンドリア膜表面あるいはミトコンドリア内のH +の移動
3)側方および膜間電子の輸送
ミトコンドリア膜表面に沿ったプロトン流は、構造化水(EZ水)によって、より高い電荷伝達速度を可能にすると考えられます。
構造化水(EZ水)については、関連記事をご参照ください ↓
ミトコンドリアの膜貫通静電局在プロトンについては、関連記事をご参照ください ↓
ミトコンドリア ナノトンネルについては、関連記事をご参照ください ↓
まとめ

電子 (マイナス)と陽子 (プラス)の形の電荷は、生物システムのあらゆる場所に存在します。
帯電した物質は、電場 (静止時)、磁場 (一定速度で移動するとき)、および電磁場 (速度が変化するとき)を生成します。
分子と高分子は、特定の電荷分布、すなわち静電ポテンシャル (MEP)を持っています。
MEP は、位置に依存する静電引力と、反発力による分子間の相互作用において重要な役割を果たします。
生体分子の固有 MEP に起因する電気力は、タンパク質間相互作用、酵素-基質相互作用、タンパク質-核酸相互作用、またはタンパク質-脂質相互作用など多くの分子相互作用を支配します。
細胞骨格(微小管やアクチンフィラメントなど) は、電荷とシグナル伝達を促進する構造ネットワークとみなすことができます。
ミトコンドリアは、膜電位が高いだけでなく、電気的に結合できるミトコンドリア ネットワークを形成できます。少なくとも 3 つの電荷伝達プロセスが起こる可能性があります。
てんかん活動や徐波活動(シータ波)の細胞外電場については、関連記事をご参照ください ↓
ギャップ結合と電気シナプスについては、関連記事をご参照ください ↓
コラーゲン線維の圧電性については、関連記事をご参照ください ↓
グラウンディング(アーシング)については、関連記事をご参照ください ↓


